
АТР был открыт в 1929 г. К. Фиске и И. Суббароу, а в 1930 г. В. Энгельгард показал возможность его образования в процессе переноса электронов по дыхательной цепи. В 1941 г. Ф. Липман выдвинул концепцию, рассматривающую АТР как «конвертируемую энергетическую валюту».
Почему в процессе эволюции именно АТР выпала такая роль? Для этого есть несколько причин, обусловленных свойствами данного соединения.
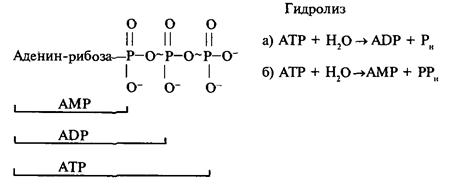
Если необходима энергия ненамного большая, чем 10 ккал/моль — по реакции Б. При необходимости энергии, значительно превышающей 10 ккал/моль, используется несколько молекул АТР в одном процессе. Иногда дополнительная энергия выделяется при сорбции АТР на ферменте.
1. Изменение свободной энергии при гидролизе фосфоангидридных связей довольно велико – около 10 ккал / моль. Когда необходима энергия меньшая или равная 10 ккал / моль, гидролиз идет по
2. Скорость неферментативного гидролиза АТР мала, т.е. молекула химически стабильна, и запасенная в ней энергия не рассеивается в виде тепла при спонтанном гидролизе. Однако замена Р на As резко повышает лабильность. Этим обстоятельством объясняется ингибиторное действие арсената на энергетический метаболизм: конкурируя с ортофосфатом, он включается вместо него в АТР, а образовавшееся соединение подвергается спонтанному гидролизу.
3. Малые размеры молекулы АТР позволяют ей свободно проникать в различные участки клетки, в то же время цитоплазматическая мембрана для нее непроницаема, следовательно, «утечка» АТР не происходит.
4. «Выбор» АТР как нуклеотида был вызван, по-видимому, необходимостью взаимодействия с белками, так как взаимодействие белков с моно- и полинуклеотидами лежит в основе жизнедеятельности.
5. «Выбор» в качестве пуриновой части молекулы аденозина, вероятно, обусловлен его промежуточными электроннодонорными и акцепторными свойствами, что обеспечивает взаимодействие с широким кругом партнеров. Кроме того, среди азотистых оснований аденин наиболее устойчив к действию ультрафиолета, что могло иметь значение на ранних этапах формирования живых систем.
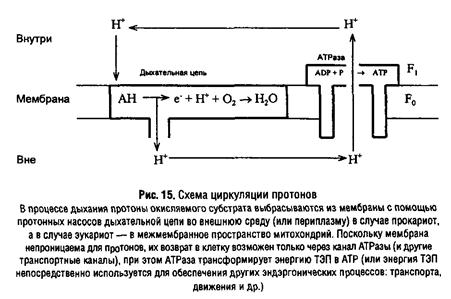
При описании механизма образования АТР путем мембранного фосфорилирования в настоящее время общепринятой является хемиосмотическая теория сопряжения окисления и фосфорилирования, предложенная П. Митчеллом в 1961 г. Согласно этой теории в «сопрягающих» мембранах локализованы два типа систем, способных к транслокации протонов: электрон-транспортная цепь и Н+-АТРаза, координированная работа которых приводит к формированию трансмембранной разности электрохимического потенциала протонов, – а затем АТР. Таким образом, первичной формой запасания энергии при дыхании является ТЭП.
Количество энергии, запасенной в форме ТЭП, прямо пропорционально количеству транслоцированных протонов: AG – пДцн+ и складывается из двух составляющих: химической и электрической:
![]()
где 2,3RT/ F = Z = 59 мВ при 25°С;
Др – протондвижущая сила.
Для образования АТР необходима AG около 250 мВ. Примерно такая величина ТЭП и создается на мембранах митохондрий и прокариотических клеток, хотя вклад каждой из составляющих различен. Например, у ацидофильных бактерий ТЭП практически полностью состоит из ЛрН, а у алкалофилов – из Л<р.
Важно отметить, что АТРазный комплекс может не только утилизировать ТЭП с образованием АТР, но и формировать его за счет гидролиза АТР, осуществляя таким образом взаимное превращение этих двух форм энергии.
Прочие статьи:
Позвоночный столб
Позвоночный столб (рис. 3, 4) — настоящая основа скелета, опора всего организма. Конструкция позвоночного столба позволяет ему, сохраняя гибкость и подвижность, выдерживать ту же нагрузку, которую может выдержать в 18 раз более толстый бе ...
Полиплоидия и отдаленная гибридизация растений
У растений одну из форм наследственной изменчивости представляет полиплоидия. Многие из культурных растений (по сравнению с родственными дикими видами) полиплоидны. К числу их относятся пшеница, картофель, некоторые сорта сахарной свеклы. ...
Новый вид смоляного удильщика Linophryne Digitopogon из
Юго- восточной атлантики
Семейство смоляных удильщиков Linophrynidae обединяет роды- Photocorynus Regan, 1926 и Borophryne Regan, 1925ю. От других глубоководных удильщиков подотряда Ceratioidei самки линофринид отличаются наличием 3(очень редко 2 или 4) лучей в с ...